




CONTRIBUTION A L’ETUDE DE LA PESTE PORCINE AFRICAINE EN REPUBLIQUE DEMOCRATIQUE DU CONGO: APPROCHES EPIDEMIOLOGIQUES ET VIROLOGIQUES.
MULUMBA-MFUMU KAZADI Léopold
THE SE PRESENTEE EN VUE DE L’OBTENTION DU GRADE DE
DOCTEUR EN SCIENCES VETERINAIRES
ANNEE ACADEMIQUE 2015-2016
Résumé
Contexteet justification de la recherche
A partir de la fin des années 1980 et pendant les années 1990, une maladie fatale, mais insidieuse apparut dans les exploitations porcines des zones annexes de la Ville de Kinshasa et ses environs qui dépendent des provinces voisines du Bas-Congo et de Bandundu. Cette maladie qui affectait surtout les porcs à partir de l’âge de 5 mois, tuait entre un et cinq sujets par semaine et ce, jusqu’à l’extermination de tout l’effectif d’une porcherie. Cliniquement, la toux, la dyspnée, la fièvre, l’anorexie, de discrets piquetés hémorragiques qui pouvaient être confondus avec de la saleté cutanée avec parfois la présence de vésicules sur l’abdomen autour des glandes mammaires étaient observés. En plus de ces observations épidémiologiques et cliniques, à l’Est de Kinshasa, il avait été constaté que les exploitations porcines d’où partaient ces alertes étaient pour la plupart situées le long de la rivière Ndjili, petit affluent du fleuve Congo, qui tire sa source dans la province voisine du Bas-Congo.
Vers avril et mai 1984, les informations reçues des mêmes zones, notamment du village Kingantoko, avaient fait état de l’occurrence d’une maladie qui, après avoir tué, tous les porcs dans une ferme porcine commerciale d’une vingtaine des porcs se serait propagée dans tout le village d’où elle avait frappé même les porcs indigènes errants, ne laissant que de rares survivants. Officiellement, y compris même par le passé, aucune déclaration de foyer
de la peste porcine africaine n’avait été faite dans cette partie de la RDC (Figure 1). Dans le même temps, des alertes similaires provenaient de l’Ouest de Kinshasa, plus précisément de Lutendele, une vallée abritant plusieurs autres exploitations porcines, non loin de la rive gauche du fleuve Congo
L’augmentation des pertes associées au syndrome décrit ci-dessus, le malentendu et la confusion entretenus par les professionnels de l’élevage sur le terrain autour du diagnostic, en pointant du doigt la maladie du Rouget, malgré l’échec du traitement avec des antibiotiques à large spectre et, le manque d’une unité spécifique de diagnostic au sein du laboratoire national de référence constituent des faits qui ont justifié cette recherche. Le but
de ce travail était de permettre une maitrise de la situation de la peste porcine Africaine en RDC à travers des enquêtes de terrain et des analyses de laboratoire.
Ainsi, six études de terrain ont été menées pouvant être regroupées en quatre thématiques :
• la confirmation ou infirmation des cas suspects de peste porcine Africaine dans les zones
à problèmes (une étude) ;
• l’étude de la prévalence, la vérification de l’état d’endémicité et l’évaluation de tests de
diagnostic utilisés sur terrain (trois études) ;
• la caractérisation moléculaire des souches du VIruS de la peste porcine africaine et
l’évaluation de leur distribution (une étude) ;
• le testage de l’efficacité d’un candidat vaccin pour la protection des porcs natifs de
l’Afrique (une étude).
Stratégie et méthodologie
Pour les enquêtes de terrain, les zones concernées par les investigations ont été choisies en fonction de l’information épidémiologique disponible, leur lien commercial avec la ville ou province de Kinshasa, leur importance en cheptel porcin, la présence d’une interface entre porcs domestiques et porcs sauvages (interactions) ainsi que leur accessibilité. C’est ainsi que les provinces de Bas-Congo, Equateur, Katanga, Orientale, Kasaï Oriental et Maniema ont été sélectionnées. Parmi ces provinces, celles qui ont été choisies en raison de la présence d’une interface entre les porcs domestiques et porcs sauvages étaient : le Bas-Congo, l’Equateur et le Kasaï Oriental.
Pour des raisons logistiques, les données épidémiologiques et les prélèvements des échantillons ont été effectués en deux périodes (1997 à 2006 et de 2005 à 2012). Les données de terrain et les prélèvements étaient enregistrés sur un fichier Excel renseignant l’origine, le lieu de chaque prélèvement, la date et la nature du prélèvement, les coordonnées géographiques (longitude et latitude), les facteurs de risque et les résultats de laboratoire.
S’agissant des facteurs de risque, les principaux étaient les suivants : l’espèce (porc domestique ou sauvage), l’âge, le poids, la race, le sexe, le système d’élevage, la présence et le type d’ ectoparasites trouvés sur la peau, les lésions hémorragiques, l’état de nutrition et la distance avec la ferme la plus proche. D’autres observations étaient notées telles que, par exemple, la température pendant le transport vers le laboratoire. Les porcs adultes inclus
dans l’étude étaient de toute race et âgés de plus de cinq mois. Au niveau de laboratoire, les méthodes suivantes ont été mises en oeuvre: l’identification des tiques, la détection des anticorps dirigés contre le virus de la peste porcine Africaine (PPA), la détection des antigènes et la détection de l’ADN du virus de la PPA, le séquençage et la construction des phylogrammes. En outre, un candidat vaccin atténué a également été testé et les outils statistiques adaptés ont été utilisés.
Pour la détection des anticorps spécifiques (Ac) dirigés contre le VIruS de la PPA et l’évaluation des. caractéristiques des tests sérologiques les plus utilisés sur terrain, trois formats d’ELISA ont été évalués. C’est ainsi que pour la sensibilisation des plaques en rapport avec lesdits formats, trois types des protéines étaient utilisés comme antigènes (Ag), à savoir:
• la protéine totale et cytosoluble p72 (cp72), utilisée dans un test ELISA indirect
approuvé comme test de référence par l’Organisation mondiale pour la Santé Animale
(OIE) et la FAO ;
• la forme recombinante de la même protéine p72 (rp72) utilisée dans un test ELISA de
compétition;
• la forme recombinante de la protéine p30 (rp30) du même virus aussi utilisée dans un test ELISA indirect.
Les tests de confirmation des résultats révélés positifs (douteux) consistaient en deux tests d’ «immunoblott » (IB) basés sur la p72 et la p30 et un test d’immunohistochimie (IRC).
Pour l’estimation relative au test de référence de la sensibilité (Se) et de la spécificité (Sp) des tests ELISA basés sur la rp72 et la rp30, quelques outils statistiques ont été utilisés dont le calcul du coefficient Kappa, la courbe ROC (Receiver Operating Characteristicï, l’estimation par noyau (Kernel density) et le test de Chi2.
Les tiques prélevées sur les porcs domestiques et sauvages étaient identifiées par scannage grâce à un microscope électronique adapté (SEM, Scanning Electron Microscope) et un microscope stéréoscopique Olympus SZX16. Cette identification a été complétée par l’exploitation d’une clé taxonomique.
Pour la caractérisation moléculaire des souches circulantes, l’ADN viral a été extrait au moyen d’un kit QIAGEN (protocole préconisé pour le sang et les autres tissus). Trois régions cibles du génome du virus de la PPA comprenant les gènes B646L, K183L et B602L (CVR) ont été amplifiés par PCR (Aguero et al., 2003 ; Bastos et al., 2003, Nix, et al., 2006).
Les amplicons ont été purifiés en utilisant le kit Wizard ® SV Gel et le kit PCR Clean Up System, en respectant le protocole préconisé par le fabricant (Promega Corporation, USA).
Les arbres phylogéniques ont été construits sur base des logiciels FASTA et MEGA (Tamura et al., 2013). Les séquences des nucléotides de tous les isolats collectés dans cette étude ont été fournies par LGC Genomics (Berlin, Allemagne) chez qui les ampli cons purifiés avaient
été soumis.
Pour la protection des porcs indigènes contre la PPA, le candidat vaccin utilisé est une souche naturellement atténuée OURT88/3, isolée sur une tique Ornithodoros au Portugal.
Huit porcs villageois ont été vaccinés avec cette souche et inoculés successivement par les souches ‘virulentes de génotype I, OURT88/l provenant du Portugal et DRC085/l0, provenant de la RDC. Chaque étape d’infection était séparée de l’autre par 21 jours d’intervalle. Un score clinique était établi et un suivi sérologique était réalisé successivement aux jours 3 (J3), 6, 14 et 21 post-inoculation (Pi). La réponse immune ainsi que la charge virale étaient suivies grâce à une PCR quantitative (qPCR). Comme témoins, 7 porcs non vaccinés ont été utilisés lors du premier challenge et, 6 porcs lors du deuxième. Les porcs vaccinés qui ont survécu après les challenges ont été sacrifiés, en adéquation avec les règles éthiques, au 21eme jour après le deuxième challenge, soit au 63eme jour de l’expérimentation.
L’essai était autorisé par les Services vétérinaires officiels de la Ville de Kinshasa.
Résultats de la recherche
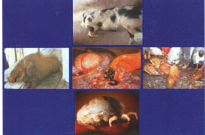
1. Par rapport aux incertitudes et confusions à propos du diagnostic de la maladie qui était à la base des pertes enregistrées à partir des années 1998-90 dans les exploitations porcines à Kinshasa et dans les environs, au lieu du Rouget, c’est la PPA qui a été confirmée. Les Ac dirigés contre le virus de la PPA ont été détectés par les tests ELISA et confirmés par les tests d’IB et d’IHC. De plus, l’ADN du virus de la PPA a été détecté par PCR dans
des échantillons de tissus prélevés dans toutes les localités où les foyers avaient été suspectés. La caractérisation moléculaire des souches impliquées a permis de savoir qu’elles appartenaient toutes au génotype I, dans un même taxon que les souches du Nigeria [Nig. 98/99] et du Ghana [Gha] (Mulumba-Mfumu et al., 2013). Nous avons déduit que la confusion du diagnostic était plus liée à la faible virulence des souches impliquées; en effet, la PPA a toujours été abusivement considérée comme une maladie exclusivement hémorragique suivie de 100% morbidité et 100% de mortalité. Ce qui n’est pas le cas dans les zones endémiques de l’Afrique sub-Saharienne. Cette croyance empirique a souvent retardé la riposte et le contrôle de la part des autorités vétérinaires et a contribué à la persistance et l’endémicité de la PPA en Afrique.
2. Concernant l’étude de la prévalence, la vérification de l’état d’endémicité et l’évaluation de tests de diagnostic utilisés sur terrain :
a) L’endémicité de la maladie a été établie, du fait de la détection des anticorps dirigés contre le virus de la PPA et des antigènes de celui-ci, autant chez les porcs domestiques apparemment sains que chez les Potamochères (Figure 5). L’évaluation.de la présence des anticorps chez les sujets apparemment sains a révélé une prévalence apparente de 27 %.
b) Concernant l’épidémiologie de la maladie, deux approches ont été envisagées lors des investigations menées à l’interface où les interactions entre les porcs domestiques et les suidés sauvages étaient possibles, notamment dans les provinces de l’Equateur (Mbandaka et Boende) et du Kasaï-Oriental (Kole). La première approche visait la confirmation de la circulation de la maladie chez les suidés sauvages et la deuxième visait la détection et prélèvement des tiques Ornithodoros dans l’écosystème de la forêt où vivent les potamochères. La première approche a permis la détection
d’anticorps dirigés contre le virus de la PPA dans les sérums prélevés de potamochères de la forêt de Lingonda (2°.35′ N, 18 0.55’E), en Province de
l’Equateur et la détection des antigènes dans les tissus lymphoïdes de potamochères de la savane en provenance de Kole (3 0.28’S, 22 °.28’E) et de Borna (5° 83 S, 13°0TE), respectivement dans les Provinces du Kasaï Oriental et du Bas-Congo. Par rapport avec la deuxième approche, aucune tique Ornithodoros n’a pu être identifiée. Ceci pourrait être mis en relation avec la méthode utilisée. En effet, plutôt que d’aller chercher les tiques dans les terriers des potamochères (difficiles à repérer dans une forêt inondée), les tiques avaient été prélevées sur les corps des suidés (domestiques
et sauvages). Par contre, parmi les tiques prélevées, plusieurs variétés des tiques ont été identifiées telles que les genres Ixodes, Amblyoma, et Parmi les Rhipicephalus, une nouvelle espèce a été découverte, non encore décrite dans la littérature et celle-ci a été nommée « Rhipicephalus congo lensis »
(Apanaskevich et al., 2013). Par ailleurs, nous n’avons pas pu amplifier de l’ADN du virus de la PPA à partir d’un broyat de toutes les tiques prélevées y compris celles provenant des bêtes séropositives (données non détaillées dans cette thèse).
c) L’évaluation des caractéristiques des tests sérologiques utilisés sur terrain a révélé, dans nos conditions de laboratoire et de terrain, une sensibilité (Se) relative de 93.48 % (95 % IC: 82.l 0-98.63) et une spécificité (Sp) relative de 87.98 % (95 % IC : 83.62-91.53) pour l’ELISA de compétition utilisant la protéine recombinant p72 (rp72). En outre, sur base des trois études indépendantes menées dans différentes zones et systèmes d’élevage, la prévalence apparente en anticorps a varié de 20 à 27 %. Par ailleurs, en prenant en considération les données de l’étude dans laquelle les autres tests de diagnostic ont été réalisés (3 tests ELISA susmentionnés et test PCR), le nombre de porcs positifs confirmés par au moins un autre test de diagnostic était de 55/328 (17%), ce qui constitue une bonne estimation de la prévalence dans la région et qui est très proche de 16,9 % noté au Sénégal par le passé (Etter et al., 20 Il).
3. Concernant les souches circulantes du virus de la PPA et leur distribution, une étude a été menée sur une période de sept années (2005 à 2012). Un séquençage basé sur trois gènes choisis comme marqueurs (B646L, K183L, B602L CVR) a permis de mettre en évidence, dans la zone d’étude, la circulation de trois génotypes p72 (I, XI, et XIV) et 19 variants. La co-circulation de différents génotypes en un même endroit (même foyer) a
également été révélée. Ceci avait déjà été démontré auparavant (Carrnina G. et al., 2010 ; Bastos, 2004). En outre, deux autres observations originales ont pu être faites, à savoir: la co-infection d’un même porc par deux variants du virus de la PPA, au cours d’un même foyer et dans une même ferme ainsi que la présence de répétions en tandems de tétramères d’acides aminés chez des isolats appartenant à différents génotypes. Ce dernier constat fait après analyse des séquences du « locus hypervariable » du génome viral (B602L-CVR) incrimine soit une homoplasie, soit une possible recombinaison génétique de plusieurs souches circulantes dans un foyer; hypothèse déjà posée dans d’autres études antérieures in vivo (Smith, 1976). Cette étude a dans le même temps aussi démontré que le génotype 1 était de loin le plus dominant dans la zone d’étude, avec 40 séquences sur 62 (64,5 %), suivi du génotype IX avec 20 séquences sur 62 (32,3%) et enfin le génotype XIV avec 2 séquences sur 62 (3,2%).
4. Concernant les perspectives de protection des porcs natifs de l’Afrique contre la PPA, un candidat vaccin basé sur une souche naturellement atténuée, OURT88/3, isolée sur une tique Ornithodoros au Portugal a été testé. Ce vaccin a été inoculé à 8 porcs natifs de l’Afrique. Les sujets inoculées ont ensuite été challengés deux fois à 21 jours d’intervalle avec deux souches virulentes, OURT8811 et DRC85110 108 provenant respectivement du Portugal et de la RDC. A la fin de cette expérimentation, 50% des porcs soit 4 sur 8 ont été protégés, un porc était mort d’une autre cause et, 3 porcs ont été tués par les souches du virus utilisées pour le challenge. Six porcs sur six, soit 100% et, six sur sept, soit 86%,
utilisés respectivement comme animaux témoins dans les challenges 1et 2 sont morts (Mulumba-Mfumu et al., 2015). Les sujets protégés ont présenté des titres élevés en anticorps dirigés contre le virus de la PPA (test ELISA) et étaient négatifs au test de PCR quantitative (qPCR). Les porcs protégés ont été euthanasiés selon les normes éthiques au 21ième jour après le deuxième challenge, soit au 63 ième jour de l’expérimentation. Ces
résultats indiquent que l’espoir de trouver un vaccin efficace contre le virus de la PPA pourrait venir de souches naturellement atténuées (Sanchez-Vizcaino, 2012).



